Meiotic drive can explain enigmatic observations in both centromere and karyotype evolution. Centromeres are the sites where kinetochores assemble and are essential for eukaryotic cell division. Although this crucial function is highly conserved, the repetitive DNA at the centromere and the centromere binding proteins have evolved rapidly. This paradox can be explained if we consider centromeres as selfish elements competing (between homologous chromosomes) for segregation to the egg in meiosis I. This form of meiotic drive, known as centromere drive, is expected to contribute to speciation because populations with different centromeres and incompatible centromere binding proteins will become reproductively isolated (Henikoff et al. 2001).
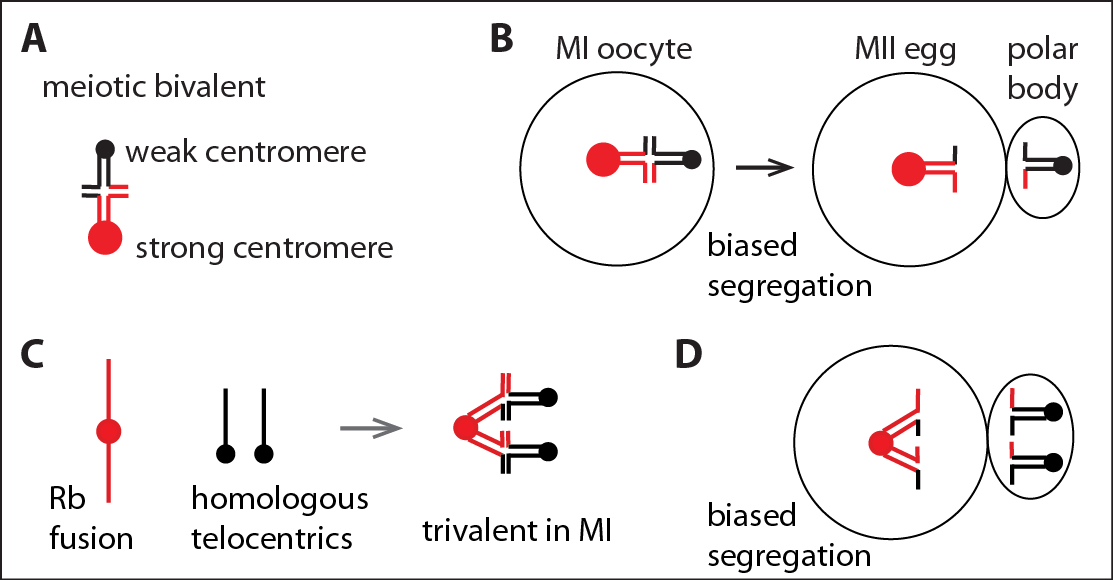 |
Meiotic drive: preferential chromosome segregation in meiosis I. (A) A meiotic bivalent with unequal centromeres. (B) Biased segregation with the stronger centromere remaining in the egg. (C) Rb fusions pair with the homologous telocentric chromosomes to form trivalents in MI. (D) Biased segregation with the Rb fusion remaining in the egg, which can lead to fixation of Rb fusions and karyotype change from mostly telocentric to mostly metacentric. |
The implications of meiotic drive for karyotype evolution are clear in the context of Robertsonian (Rb) fusions: common chromosomal rearrangements in mammals in which two telocentric chromosomes (centromere at one end) join at their centromeres to form a single metacentric (centromere internal). Biased segregation of Rb fusions can explain massive karyotype change on short evolutionary timescales in mammals (White et al. 2010) and the striking bimodal distribution of karyotypes in many species: karyotypes that are either primarily telocentric or primarily metacentric are common, but “mixed” karyotypes are rare (Pardo-Manuel de Villena and Sapienza, 2001, Molina et al. 2014). Furthermore, meiotic drive acting on chromosomal rearrangements, such as Rb fusions, can lead to speciation because hybrid individuals would be infertile (White 1968). We provided the first cell biological evidence linking differences in centromere-microtubule interactions to biased segregation of Rb fusion chromosomes. We also showed, using both laboratory models and mice from natural populations, that natural variation in centromere strength can explain why Rb fusions preferentially segregate to the egg in some populations, leading to karyotype change, and not others (Chmátal et al. 2014).
 |
Our model system for meiotic drive of Rb fusion chromosomes. A fusion between mouse chromosomes 6 and 16 pairs with the homologous telocentric chromosomes to form a trivalent in female meiosis I. See Chmátal et al. 2014. |
We have established experimentally tractable mouse model systems (Chmátal et al. 2014, Iwata-Otsubo et al. 2017, Akera et al. 2019) to test mechanistic models for how chromosomes can “win” in meiosis to increase their transmission through the female germline. Our findings are providing the first cell biological insights into the cell biology of meiotic drive in animals. We showed that selfish centromeres preferentially segregate to the egg through interactions with an asymmetric meiotic spindle (Akera et al. 2017). Exploiting centromere divergence between species, we showed that distinct mechanisms can modulate a common molecular pathway that regulates interactions between centromeres and spindle MTs to achieve biased segregation (Akera et al. 2019). We established a conceptual framework for both drive by selfish centromere DNA sequences and suppression by rapidly evolving centromere proteins, based on parallel kinetochore and heterochromatin pathways for recruiting drive effectors (Kumon et al. 2021). With broad consequences for reproductive biology, chromosome evolution, and genetics, this project represents a unique contribution to the study of chromosome segregation and to evolutionary cell biology.
 |
Model system for centromere drive. A mouse oocyte from a hybrid female produced by crossing strains with different centromeres. None of the chromosomes align at a metaphase plate because centromeres inherited from the two parents interact differently with spindle microtubules, ultimately lead to preferential segregation of centromeres from one parent to the egg. The image shows metaphase of meiosis I, with chromosomes (green) and centromeres (red) labeled. Both bivalents and trivalents are present. See Chmátal et al. 2014, Iwata-Otsubo et al. 2017. |
 |
Meiotic spindle asymmetry. Signals from the polarized cell cortex (green) in mouse oocytes regulate microtubule tyrosination (white) to generate spindle asymmetry in meiosis I. This asymmetry can be exploited by selfish centromeres to bias their transmission to the egg. See Akera et al. 2017. |